Alessandro M Lazdins, Claire E Miller, Mark A Webber and Christopher M Thomas,
Institute of Microbiology and Infection, University of Birmingham, UK
Antibiotic resistant infections have been recognized as a major global health problem that is intensifying due to the lack of concrete novel approaches to fight them. Plasmids are recognized as one of the major contributors to the rapid worldwide spread of resistance genes. However, few strategies that specifically target plasmids are currently being pursued. Our work is developing ways to displace plasmids from their hosts using incompatibility functions without disrupting cell viability by neutralizing plasmid-encoded toxin/anti-toxin systems. This approach is giving promising in vitro results, but needs further development and recognition as a viable option to fight the problem of resistance.
The current widespread occurrence of multidrug-resistant (MDR) bacteria has become a major burden on national health services, with death rates due to bacterial infections on the rise. One of the factors that heavily contributes to the problem is how rapidly novel and existing resistance determinants move among bacterial hosts. Many globally successful strains of pathogens have been able to become widely disseminated as a result of resistance genes acquired on mobile genetic elements (MGEs) (1). This rapid spread is in part due to the extensive movement of people, goods and animals for example, all of which allow for the interaction between different bacterial strains and their MGEs, resulting in a globalized gene pool (2).
Plasmids
Joshua Lederberg first coined the term “plasmid” in 1952 (3), and since their discovery as “extra-chromosomal hereditary determinants”, studies have highlighted their role in horizontal gene transfer (HGT) and usefulness as tools in molecular biology. Much of the early work on plasmids focused on the observation that resistance to antibiotics seemed to transfer from one strain to another, leading to the description of R-factors (4, 5). However, in the early 1970s, Stanley Falkow, Stanley Cohen, Herbert Boyer, Donald Helinski, Charles Brinton and several others developed the concept of using plasmids as tools for gene cloning. The discovery that plasmids could be efficiently used as cloning vectors led to an increased interest in understanding plasmid biology, as well as stimulating new types of biotechnology.
Plasmids are typically circular (but also less commonly linear) double-stranded DNA molecules that are able to independently control their multiplication and stable inheritance from generation to generation in their bacterial hosts (2). In addition, many plasmids can transfer from one bacterium to another, the most sophisticated mechanism being by conjugation in which the plasmid carries genes that can create a bridge between bacteria through which a copy of the plasmid can move (Fig 1a). Some plasmids are not able to do this on their own, but can use the bridge made by other plasmids to transfer themselves. The set of genes for multiplication, stable inheritance and transfer are called the plasmid backbone or core. Plasmids can also pick up a variable cargo of other genes that can help their host bacterium grow or survive in different environments – the plasmid spreads them between bacteria and if a survival advantage is gained due to the carried genes on the plasmid there will be positive selection for its carriage. Amongst the favourable traits carried on plasmids, genes conferring antibiotic resistance are of particular concern in the spread of drug-resistant infections (6).
Although all plasmids basically function in similar ways, what makes targeting plasmids difficult is that the genes and proteins they need for multiplication and stable inheritance are highly diverse making it unlikely to find a single compound that will block them all.
Dominant plasmids
The role of plasmids, particularly those able to transfer autonomously (conjugative), in the spread of antibiotic resistance was quickly established. But it was not until techniques like DNA sequencing became commonplace that the extent of the role plasmids play in the dissemination and evolution of resistance traits became clear (7). Genetically identical plasmids were identified globally in different, completely unrelated strains of bacteria. Genes such as NDM-1 conferring resistance to β-lactam antibiotics, and other essential antibiotics, have been observed on dominant plasmid types that are known to have been in circulation for over 50 years. For example, resistance determinants on one type of plasmid isolated from outbreaks of disease caused by E. coli in Canada were also identified only a few years later on different plasmid backbones in outbreaks of K. pneumoniae infection in Sweden . It appears that past antibiotic usage has selected certain plasmids, which are then more likely to pick up the next resistance gene that comes along.

Incompatibility and toxin/anti-toxin systems
All plasmids have ways to control their multiplication so that they do not become a burden to their host bacterium. Closely related plasmids that use the same or similar multiplication genes will therefore compete with each other and cannot be stably inherited together. This is called plasmid “incompatibility” and has been historically used to classify plasmids into different incompatibility groups – plasmids that compete with each other are classified into the same incompatibility group (11). As DNA sequencing technologies developed it became clear that groups of incompatible plasmids shared key parts of their backbones and could be identified by PCR methods . Another class of evolutionary adaptation to ensure stable inheritance inside a host are the toxin/anti-toxin (TA) systems. Many plasmids carry such TA systems, which are usually composed of a stable toxin and an unstable anti-toxin. This means that if the plasmid is lost, the stable toxin outlives the unstable anti-toxin, resulting in death or reduced viability of the plasmid-free cell, and thus maintenance of the plasmid within the overall population (14).
Plasmid curing
As the problem of antibiotic resistance grows, and the number of new antibiotics fails to provide a solution, novel approaches are required to tackle the issue. Since the initial discovery of plasmids, efforts have been made to eliminate, or “cure”, them from their hosts in order to better understand the biological role that these elements have. Early attempts to eliminate plasmids by Salisbury et al. (15) involved stressing the host either by growing the bacterium at higher than optimal temperatures, or by adding chemical compounds such as acridine orange, sodium dodecyl sulfate (SDS) or ethidium bromide. These interventions were only successful for some plasmids and generally also resulted in mutations and damage to the host. Approaches such as these are also not amenable to use in curing plasmids beyond the laboratory.
As more was learned about plasmid replication in the 1970s and with the development of gene cloning, plasmid regions that specifically interfered with stable inheritance were discovered. Different groups, including Hynes et al. (16), developed techniques that relied on identifying the incompatibility groups of the target plasmids and then introducing plasmids with the same incompatibility group to induce curing. This technique was refined by Stolt et al. (17), who cloned only the specific regions that determine incompatibility onto a vector, and showed that this could cure a plasmid carried by Mycobacterium fortuitum. Uraji et al. (18) refined the work of Stolt et al. (1996), by cloning the replication functions to induce incompatibility onto a vector carrying a gene that could be induced to self-destruct. This allowed for the selection of completely plasmid-free strains.
Although the mentioned studies demonstrated the feasibility of using incompatibility to stably cure plasmids from bacterial populations, using this, as an alternative approach to deal with increasingly antibiotic resistance strains was first truly conceptualized in a patent application filed by Filutowicz (19).
Denap et al. (20) took the concept of incompatibility and used it as a rationale to test small chemicals that specifically target functions required for plasmid replication. They used aminoglycosides, which are known to target RNA, and tested their ability to block the replication of plasmids that use RNA as control elements during replication. Their studies showed that when using these drugs a bacterial population could be re-sensitized. However, using antibiotics to try and reduce the problem of antibiotic resistance is not an optimal strategy and many plasmids carry aminoglycoside resistance genes. Nevertheless, it brought forward the concept that plasmids could be a good target for new therapeutic strategies, an idea which was supported by Latha et al. (21) who isolated plant extracts with anti-plasmid properties.
More recently, several groups including ours have looked at ways to specifically target plasmids that carry antibiotic resistance using genetic techniques. Bikard et al. (22) and Ji et al. (23) used CRISPR-Cas to design target-specific systems delivered either by bacteriophages (22) or conjugative plasmids (23). These approaches highlight how genetic techniques can be used instead of chemical compounds as antimicrobials and warrants further development alongside approaches such as phage therapy.
pCURE
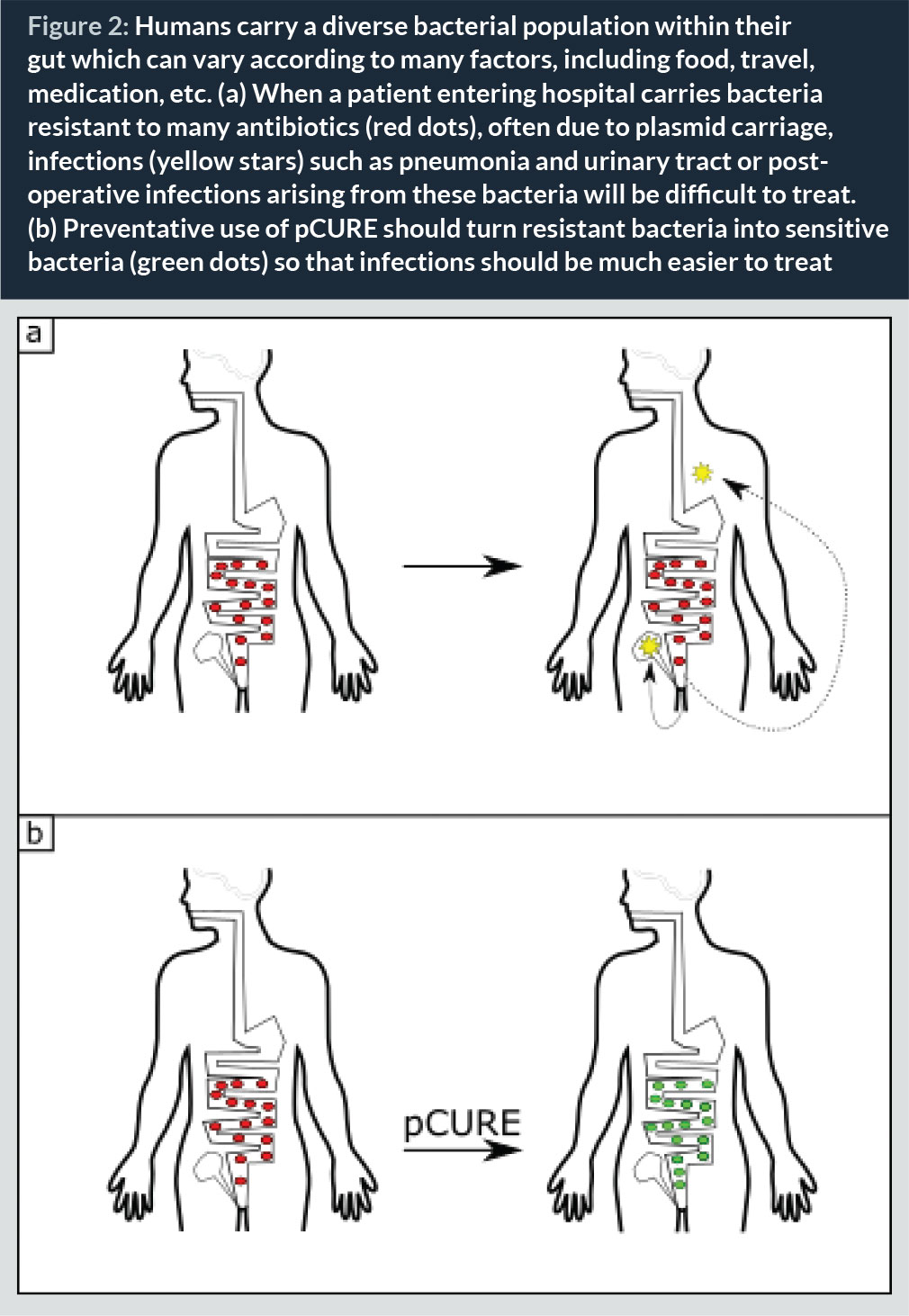
The presence of large antibiotic resistance plasmids in bacterial populations is the major determinant of poor clinical outcome during hospital-acquired infections (HAI). Vulnerable or immunocompromised patients commonly suffer from HAIs when bacteria that are normally carried in the gut or on the skin infect wound sites following surgery or form biofilms on indwelling devices such as ventilators and catheters leading to pneumonia or UTIs (Fig. 2). Due to the facility with which plasmids can transfer among bacteria, including gut bacteria, developing a system whose purpose is to displace antibiotic resistance plasmids from particular environments could be a good approach to reduce the burden of antibiotic resistance.
The pCURE system was developed in the Thomas laboratory by combining functions to block replication of a target plasmid whilst also neutralizing any TA system it carries (Fig. 1b). These “curing functions” are cloned into a different vector that is not itself incompatible with (and therefore blocked by) the target plasmid(s). The combination of targeting multiple replicons and neutralizing TA systems makes our approach unique and essential for applications in environments where one would not want to disrupt the normal ecological balance. For example, it could provide a clear advantage over antibiotic treatment, which can often disrupt the gut microbiota and can result in overgrowth of resistant pathogens such as C. difficile. Hale et al. (24) showed that the pCURE system could be engineered and applied to target different types of plasmids and that it cured at very high efficiency. Although useful in an in vitro setting as shown by several groups who used the system to cure important resistance plasmids (25, 26), the pCURE described by Hale et al. (24) could not be used in more complex environments because it is not self-transmissible.
Currently work in our group is being undertaken to develop a pCURE system that is self-transmissible and could therefore spread among complex microbial communities, displacing antibiotic resistance plasmids as it does so (Fig. 1c). To achieve this goal we are using conjugative IncP-1 plasmids as the core of our vector because of their very broad host range (27) allowing efficient spread through complex communities. We have so far shown that these conjugative pCURE plasmids can transfer, invade and displace an IncF plasmid from a bacterial population in the absence of external selection (Lazdins et al. unpublished results). Work is currently ongoing to expand the curing potential to target a range of natural antibiotic-resistance plasmids and to optimize transfer rates.
Future work and applications for pCURE
In countries where general rates of antibiotic resistance are low (e.g., The Netherlands) it is common practice that all patients entering intensive care units (ITUs) undergo selective digestive and/or oropharyngeal decontamination (SDD and SOD) (28). These involve the administration of prophylactic antibiotics in order to try and reduce the prevalence of infections that would lead to increased mortality. Although effective in decreasing the mortality rates (29), the disruption caused to the normal flora and the possible development of further antibiotic-resistant strains is not ideal. Our vision for pCURE is that it could be used as a prophylactic measure in a similar way to SDD or SOD, given to patients who enter healthcare settings. By giving pCURE to patients at risk of developing infections upon admission to hospital, the idea is that if an infection develops, then it is more likely for it to be sensitive to standard antibiotic therapy, due to displacement of the plasmids carrying resistance (Fig. 2). This can also be applied to healthcare workers to limit possible spread of any antibiotic-resistant strains that they may be carrying.
Another area where we think that pCURE would be particularly useful is in limiting the international spread of resistance. There are places where the number of strains carrying plasmids with multiple antibiotic resistance genes is higher than others (Southeast Asia vs. Scandinavia for example) and studies have demonstrated that individuals travelling to places with higher occurrence will quickly be colonized by resistance strains even without becoming ill and these strains can remain in their flora for extended periods (30). Therefore, upon returning from places with a high risk of colonization, travellers could take a course of pCURE to eradicate any problematic resistance genes they have acquired. For this vision to be realised several steps need to be taken to ensure that pCURE becomes a cost-efficient, adaptable and, most importantly, safe system to use in human medicine.
Bacterial conjugation in the gastrointestinal tract (GIT) of humans is poorly understood, and although there is evidence that conjugation is an important factor influencing HGT in the GIT (See (31)) for a current review), more work will be needed to fully understand the extent of pCURE spread once administered. These studies will require the use of animal models or complex in vitro gut models. Once these studies are done, more work can be undertaken to optimize pCURE as a transfer agent, but also to find the best way to administer it. We are currently taking inspiration from probiotics and faecal transplant treatments, where in both cases live bacteria are successfully delivered to the gastrointestinal tract.
Safety considerations are essential when establishing how best to administer pCURE and track how it spreads. Plasmids have the ability to pick up resistance genes extremely easily as well as having the ability to integrate into bacterial-host chromosomes. To ensure safety and avoid fears of releasing genetically modified elements into nature, we would need to prevent the acquisition of resistance genes by the pCURE vector as well as the potential for chromosomal integration by including genetic safeguards in our system. These would include systems that specifically block the ability of plasmids to acquire extra resistance genes as well introducing specific self-destruct mechanisms to avoid pCURE from working outside of the body.
Extensive discussions will also be needed with safety governing bodies (for example, the FDA or EMA) regarding the classification and requirements for the usage of pCURE. At present, pCURE sits somewhere between a food, a drug and a biologic so specific requirements for trials and approval in human usage will need to be clarified. Alongside this clarification, studies on how the public would perceive pCURE as a product are essential. A lot is known about the reticence that people have for using genetically modified organisms (GMOs) (32, 33), but would this reticence change in the face of the dangers of antibiotic resistance?
Conclusion
Even though plasmids are the major reason for the development and spread of resistance among naïve bacterial populations, very little attention is being given to these elements as targets for combating the maintenance and spread of antibiotic resistance. The knowledge and technology to develop plasmid-curing technologies is accessible, and many groups have used a range of approaches to achieve this goal in vitro. However, the jump to using these kinds of systems in more complex environments has yet to be realized. We believe that the practical value of pCURE should be explored as an efficient way to cure antibiotic-resistance plasmids from strains colonizing the GIT in a way that would not disrupt the normal flora, resulting in a reduced clinical burden if infection were to manifest itself.
Biographies
Dr Claire Miller gained her PhD in Bacterial Genetics from the University of Leicester, followed by a three year post-doctoral research position in the same laboratory. She is currently working in the laboratory of ProfessorThomas at the University of Birmingham. She joined the laboratory on a Wellcome Trust ISSF Translational Award to develop plasmid curing as a method for reducing antibiotic resistance carriage in bacteria.
Dr Mark A Webber is interested in the study of the mechanisms and biology of antibiotic resistance, bacterial responses to stress and biofilm formation and has published over 50 articles in these areas since 2001. He was awarded a BBSRC David Phillips Fellowship in 2007 and took up a tenured position within the Institute for Microbiology and Infection at Birmingham at the beginning of 2012.
Professor Christopher M Thomas is Professor of Molecular Genetics in the School of Biosciences and Institute of Microbiology and Infection at the University of Birmingham. He has been passionate about bacterial plasmids as systems ever since his DPhil in Oxford with Keith Dyke and postdoc with Donald Helinski at University of California, San Diego. He led the creation of the International Society for Plasmid Biology in 2004 and set up Plasgene in 2006 based on plasmid curing technology.
To download this article please click here
” We hope that you will enjoy the 2016 edition of AMR Control.“
References
1. WHO. Antimicrobial resistance: Global report on surveillance. Geneva, Switzerland: World Health Organization; 2014.
2. Thomas CM, Nielsen KM. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat Rev Microbiol. 2005;3(9):711-21.
3. Lederberg J. Cell genetics and hereditary symbiosis. Physiol Rev. 1952;32(4):403-30.
4. Nakaya R, Nakamura A, Murata Y. Resistance transfer agents in Shigella. Biochem Biophys Res Commun. 1960;3:654-9.
5. Watanabe T. Infective heredity of multiple drug resistance in bacteria. Bacteriol Rev. 1963;27:87-115.
6. Carattoli A. Plasmids and the spread of resistance. Int J Med Microbiol. 2013;303(6-7):298-304.
7. Norman A, Hansen LH, Sorensen SJ. Conjugative plasmids: vessels of the communal gene pool. Philos Trans R Soc Lond B Biol Sci. 2009;364(1527):2275-89.
8. Woodford N, Carattoli A, Karisik E, Underwood A, Ellington MJ, Livermore DM. Complete nucleotide sequences of plasmids pEK204, pEK499, and pEK516, encoding CTX-M enzymes in three major Escherichia coli lineages from the United Kingdom, all belonging to the international O25:H4-ST131 clone. Antimicrob Agents Chemother. 2009;53(10):4472-82.
9. Boyd DA, Tyler S, Christianson S, McGeer A, Muller MP, Willey BM, et al. Complete nucleotide sequence of a 92-kilobase plasmid harboring the CTX-M-15 extended-spectrum beta-lactamase involved in an outbreak in long-term-care facilities in Toronto, Canada. Antimicrob Agents Chemother. 2004;48(10):3758-64.
10. Sandegren L, Linkevicius M, Lytsy B, Melhus A, Andersson DI. Transfer of an Escherichia coli ST131 multiresistance cassette has created a Klebsiella pneumoniae-specific plasmid associated with a major nosocomial outbreak. J Antimicrob Chemother. 2012;67(1):74-83.
11. Novick RP. Plasmid incompatibility. Microbiol Rev. 1987;51(4):381-95.
12. Carattoli A, Bertini A, Villa L, Falbo V, Hopkins KL, Threlfall EJ. Identification of plasmids by PCR-based replicon typing. J Microbiol Methods. 2005;63(3):219-28.
13. Bertini A, Poirel L, Mugnier PD, Villa L, Nordmann P, Carattoli A. Characterization and PCR-based replicon typing of resistance plasmids in Acinetobacter baumannii. Antimicrob Agents Chemother. 2010;54(10):4168-77.
14. Yamaguchi Y, Park JH, Inouye M. Toxin-antitoxin systems in bacteria and archaea. Annu Rev Genet. 2011;45:61-79.
15. Salisbury V, Hedges RW, Datta N. Two modes of “curing” transmissible bacterial plasmids. J Gen Microbiol. 1972;70(3):443-52.
16. Hynes MF, Simon R, Puhler A. The development of plasmid-free strains of Agrobacterium tumefaciens by using incompatibility with a Rhizobium meliloti plasmid to eliminate pAtC58. Plasmid. 1985;13(2):99-105.
17. Stolt P, Stoker NG. Functional definition of regions necessary for replication and incompatibility in the Mycobacterium fortuitum plasmid pAL5000. Microbiology. 1996;142 ( Pt 10):2795-802.
18. Uraji M, Suzuki K, Yoshida K. A novel plasmid curing method using incompatibility of plant pathogenic Ti plasmids in Agrobacterium tumefaciens. Genes Genet Syst. 2002;77(1):1-9.
19. Filutowicz M. Displacing a plasmid in a bacterial population. Google Patents; 2004.
20. Denap JC, Thomas JR, Musk DJ, Hergenrother PJ. Combating drug-resistant bacteria: small molecule mimics of plasmid incompatibility as antiplasmid compounds. J Am Chem Soc. 2004;126(47):15402-4.
21. Latha C, Shriram VD, Jahagirdar SS, Dhakephalkar PK, Rojatkar SR. Antiplasmid activity of 1’-acetoxychavicol acetate from Alpinia galanga against multi-drug resistant bacteria. Journal of Ethnopharmacology. 2009;123(3):522-5.
22. Bikard D, Euler CW, Jiang W, Nussenzweig PM, Goldberg GW, Duportet X, et al. Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials. Nat Biotechnol. 2014;32(11):1146-50.
23. Ji W, Lee D, Wong E, Dadlani P, Dinh D, Huang V, et al. Specific gene repression by CRISPRi system transferred through bacterial conjugation. ACS Synth Biol. 2014;3(12):929-31.
24. Hale L, Lazos O, Haines A, Thomas C. An efficient stress-free strategy to displace stable bacterial plasmids. Biotechniques. 2010;48(3):223-8.
25. Ho NK, Crandall I, Sherman PM. Identifying mechanisms by which Escherichia coli O157:H7 subverts interferon-gamma mediated signal transducer and activator of transcription-1 activation. PLoS One. 2012;7(1):e30145.
26. Kirchner M, Abuoun M, Mafura M, Bagnall M, Hunt T, Thomas C, et al. Cefotaxime resistant Escherichia coli collected from a healthy volunteer; characterisation and the effect of plasmid loss. PLoS One. 2013;8(12):e84142.
27. Popowska M, Krawczyk-Balska A. Broad-host-range IncP-1 plasmids and their resistance potential. Frontiers in Microbiology. 2013;4:44.
28. de Jonge E, Schultz MJ, Spanjaard L, Bossuyt PM, Vroom MB, Dankert J, et al. Effects of selective decontamination of digestive tract on mortality and acquisition of resistant bacteria in intensive care: a randomised controlled trial. Lancet. 2003;362(9389):1011-6.
29. Price R, MacLennan G, Glen J, Su DC. Selective digestive or oropharyngeal decontamination and topical oropharyngeal chlorhexidine for prevention of death in general intensive care: systematic review and network meta-analysis. BMJ. 2014;348:g2197.
30. Ostholm-Balkhed A, Tarnberg M, Nilsson M, Nilsson LE, Hanberger H, Hallgren A, et al. Travel-associated faecal colonization with ESBL-producing Enterobacteriaceae: incidence and risk factors. J Antimicrob Chemother. 2013;68(9):2144-53.
31. Huddleston JR. Horizontal gene transfer in the human gastrointestinal tract: potential spread of antibiotic resistance genes. Infect Drug Resist. 2014;7:167-76.
32. Savadori L, Savio S, Nicotra E, Rumiati R, Finucane M, Slovic P. Expert and public perception of risk from biotechnology. Risk Anal. 2004;24(5):1289-99.
33. Marris C. Public views on GMOs: deconstructing the myths. Stakeholders in the GMO debate often describe public opinion as irrational. But do they really understand the public? EMBO Rep. 2001;2(7):545-8.